
Glicerofosfolípido - Glycerophospholipid


Los glicerofosfolípidos o fosfoglicéridos son fosfolípidos a base de glicerol . Son el componente principal de las membranas biológicas .
Estructuras
El término glicerofosfolípido significa cualquier derivado de ácido glicerofosfórico que contiene al menos un O - acilo , o O - alquilo , o O alqu-1'-enil residuo unido al glicerol resto .
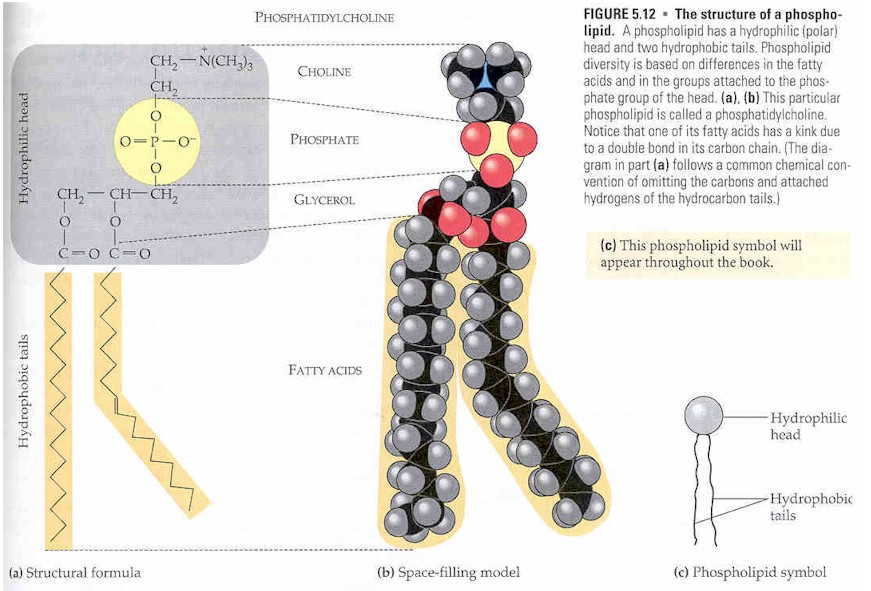
El alcohol aquí es glicerol, al que se unen dos ácidos grasos y ácido fosfórico como ésteres. Las dos cadenas de ácidos grasos unidas a la molécula de glicerol son no polares, por lo tanto, hidrófobas, mientras que las cabezas polares, que consisten principalmente en el grupo fosfato unido al tercer carbono de la molécula de glicerol, son hidrófilas. Esta doble característica conduce a la naturaleza anfipática de los glicerofosfolípidos. Por lo general, se organizan en una bicapa en membranas con las cabezas hidrófilas polares que se pegan hacia afuera al ambiente acuoso y las colas hidrófobas no polares apuntando hacia adentro. Los glicerofosfolípidos constan de varias especies diversas que suelen diferir ligeramente en estructura. La estructura más básica es un fosfatidato. Esta especie es un intermedio importante en la síntesis de muchos fosfoglicéridos. La presencia de un grupo adicional unido al fosfato permite muchos fosfoglicéridos diferentes.
Por convención, las estructuras de estos compuestos muestran los 3 átomos de carbono de glicerol verticalmente con el fosfato unido al átomo de carbono número tres (en la parte inferior). Los plasmalógenos y los fosfatidatos son ejemplos.
Nomenclatura y estereoquímica
En general, los glicerofosfolípidos usan una notación "sn", que significa numeración estereoespecífica . Cuando aparecen las letras "sn" en la nomenclatura, por convención, el grupo hidroxilo del segundo carbono del glicerol (2- sn ) está a la izquierda en una proyección de Fischer . La numeración sigue la de las proyecciones de Fischer, siendo 1- sn el carbono en la parte superior y 3- sn el inferior.
La ventaja de esta notación particular es que la configuración espacial ( D o L ) de la glicero-molécula está determinada intuitivamente por los residuos en las posiciones sn -1 y sn -3.
Por ejemplo, el ácido sn -glicero-3- fosfórico y el ácido sn -glicero-1-fosfórico son enantiómeros .
La mayoría de los aceites vegetales tienen ácidos grasos insaturados en la posición sn -2, con ácidos grasos saturados en la posición 1- sn y / o 3- sn . Las grasas animales tienen con mayor frecuencia ácidos grasos saturados en la posición 2- sn , con ácidos grasos insaturados en la posición 1- sn y / o sn3.
Ejemplos de
- Plasmalógenos
Los plasmalógenos son un tipo de fosfoglicéridos. El primer carbono del glicerol tiene una cadena de hidrocarburo unida a través de un enlace éter, no éster. Los enlaces son más resistentes al ataque químico que los enlaces éster. El segundo átomo de carbono (central) tiene un ácido graso unido por un éster. El tercer carbono se une a una etanolamina o colina por medio de un éster de fosfato. Estos compuestos son componentes clave de las membranas de músculos y nervios.
- Fosfatidatos
Los fosfatidatos son lípidos en los que los dos primeros átomos de carbono del glicerol son ésteres de ácidos grasos y el 3 es un éster de fosfato. El fosfato sirve como enlace con otro alcohol, generalmente etanolamina, colina, serina o un carbohidrato. La identidad del alcohol determina la subcategoría del fosfatidato. Hay una carga negativa en el fosfato y, en el caso de colina o serina, un ion amonio cuaternario positivo. (La serina también tiene un grupo carboxilato negativo). La presencia de cargas da una "cabeza" con una carga general. La porción de éster de fosfato ("cabeza") es hidrófila, mientras que el resto de la molécula, la "cola" de ácido graso, es hidrófoba. Estos son componentes importantes para la formación de bicapas lipídicas.
Las fosfatidiletanoaminas, las fosfatidilcolinas y otros fosfolípidos son ejemplos de fosfatidatos.
- Fosfatidilcolinas
Las fosfatidilcolinas son lecitinas . La colina es el alcohol, con un amonio cuaternario cargado positivamente, unido al fosfato, con una carga negativa. Las lecitinas están presentes en todos los organismos vivos. Una yema de huevo tiene una alta concentración de lecitinas, que son comercialmente importantes como agentes emulsionantes en productos como la mayonesa. Las lecitinas también están presentes en el tejido cerebral y nervioso.
- Otros fosfolípidos
Hay muchos otros fosfolípidos, algunos de los cuales son glicolípidos . Los glicolípidos incluyen azúcares fosfatidílicos donde el grupo funcional alcohol es parte de un carbohidrato. Los azúcares fosfatidílicos están presentes en plantas y ciertos microorganismos. Un carbohidrato es muy hidrófilo debido a la gran cantidad de grupos hidroxilo presentes.
Usos
Funciones y uso en membranas
Una de las principales funciones del glicerofosfolípido es servir como componente estructural de las membranas biológicas. Su naturaleza anfipática impulsa la formación de la estructura bicapa lipídica de las membranas. La membrana celular vista bajo el microscopio electrónico consta de dos capas identificables, o "valvas", cada una de las cuales está formada por una fila ordenada de moléculas de glicerofosfolípidos. La composición de cada capa puede variar mucho según el tipo de celda.
- Por ejemplo, en los eritrocitos humanos, el lado citosólico (el lado que mira al citosol ) de la membrana plasmática consiste principalmente en fosfatidiletanolamina , fosfatidilserina y fosfatidilinositol.
- Por el contrario, el lado exoplasmático (el lado en el exterior de la célula) consiste principalmente en fosfatidilcolina y esfingomielina , un tipo de esfingolípido .
Cada molécula de glicerofosfolípidos consta de un pequeño grupo de cabeza polar y dos largas cadenas hidrófobas . En la membrana celular, las dos capas de fosfolípidos se organizan de la siguiente manera:
- las colas hidrofóbicas se apuntan entre sí y forman un centro graso e hidrofóbico
- Los grupos de cabezas iónicas se colocan en las superficies interna y externa de la membrana celular.
Esta es una estructura estable porque los grupos de cabeza hidrófilos iónicos interactúan con los medios acuosos dentro y fuera de la célula, mientras que las colas hidrófobas maximizan las interacciones hidrófobas entre sí y se mantienen alejadas de los ambientes acuosos . El resultado general de esta estructura es la construcción de una barrera grasa entre el interior de la célula y su entorno.
Además de su función en las membranas celulares, funcionan en otros procesos celulares como la inducción y el transporte de señales. En cuanto a la señalización, proporcionan los precursores de prostanglandinas y otros leucotrienos. Es su distribución y catabolismo específicos lo que les permite llevar a cabo los procesos de respuesta biológica enumerados anteriormente. Su función como centros de almacenamiento de mensajeros secundarios en la membrana también es un factor que contribuye a su capacidad para actuar como transportadores. También influyen en la función de las proteínas. Por ejemplo, son componentes importantes de las lipoproteínas (proteínas solubles que transportan la grasa en la sangre), por lo que afectan su metabolismo y función.
Uso en emulsificación
Los glicerofosfolípidos también pueden actuar como un agente emulsionante para promover la dispersión de una sustancia en otra. A veces se utiliza en la elaboración de dulces y helados.
En el cerebro
Las membranas neurales contienen varias clases de glicerofosfolípidos que se renuevan a diferentes velocidades con respecto a su estructura y localización en diferentes células y membranas. Hay tres clases principales a saber; 1-alquil-2-acil glicerofosfolípido, 1,2-diacil glicerofosfolípido y plasmalógeno. La función principal de estas clases de glicerofosfolípidos en las membranas neurales es proporcionar estabilidad, permeabilidad y fluidez a través de alteraciones específicas en sus composiciones. La composición de glicerofosfolípidos de las membranas neurales altera enormemente su eficacia funcional. La longitud de la cadena de acilo de glicerofosfolípidos y el grado de saturación son determinantes importantes de muchas características de la membrana, incluida la formación de dominios laterales que son ricos en ácidos grasos poliinsaturados. La degradación de glicerofosfolípidos mediada por receptores por las fosfolipasas A (1), A (2), C y D da como resultado la generación de segundos mensajeros, como prostaglandinas , eicosanoides , factor activador de plaquetas y diacilglicerol. Por tanto, los fosfolípidos de la membrana neural son un reservorio de segundos mensajeros. También participan en la apoptosis, la modulación de las actividades de los transportadores y las enzimas unidas a la membrana. Se ha informado que se producen alteraciones marcadas en la composición de glicerofosfolípidos de la membrana neural en trastornos neurológicos. Estas alteraciones dan como resultado cambios en la fluidez y permeabilidad de la membrana. Estos procesos, junto con la acumulación de peróxidos de lípidos y el metabolismo energético comprometido, pueden ser responsables de la neurodegeneración observada en los trastornos neurológicos.
Metabolismo
El metabolismo de los glicerofosfolípidos es diferente en eucariotas, células tumorales y procariotas. La síntesis en procariotas implica la síntesis de glicerofosfolípidos, ácido fosfatídico y grupos de cabezas polares. La síntesis de ácido fosfatídico en eucariotas es diferente, hay dos rutas, una a la otra, hacia la fosfatidilcolina y la fosfatidiletanolamina. Los glicerofosfolípidos generalmente se metabolizan en varios pasos con diferentes intermediarios. El primer paso en este metabolismo implica la adición o transferencia de las cadenas de ácidos grasos al esqueleto de glicerol para formar el primer intermedio, el ácido lisofosfatídico (LPA). A continuación, el LPA se acila para formar el siguiente intermedio ácido fosfatídico (PA). El PA se puede desfosforilar dando lugar a la formación de diacilglicerol, que es esencial en la síntesis de fosfatidilcolina (PC). PC es una de las muchas especies de glicerofosfolípidos. En una vía llamada vía Kennedy, las cabezas polares se agregan para completar la formación de toda la estructura que consta de las regiones de la cabeza polar, las dos cadenas de ácidos grasos y el grupo fosfato unido a la columna vertebral de glicerol. En esta vía de Kennedy, la colina se convierte en CDP-colina que impulsa la transferencia de los grupos de cabezas polares para completar la formación de PC. A continuación, la PC se puede convertir en otras especies de glicerofosfolípidos como la fosfatidilserina (PS) y la fosfatidiletanolamina (PE).
Ver también
Referencias
enlaces externos
- Glicerofosfolípidos en los encabezados de materias médicas (MeSH) de la Biblioteca Nacional de Medicina de EE. UU .
- Diagrama en uca.edu
{kind=link}