
caparazón gasterópodo -Gastropod shell


1 – ombligo
2 – trenza columelar
3 – abertura
4 – columela
5 – sutura
6 – verticilo del cuerpo
7 – vértice

El caparazón de gasterópodo es parte del cuerpo de un gasterópodo o caracol, una especie de molusco . El caparazón es un exoesqueleto , que protege de los depredadores, el daño mecánico y la deshidratación, pero también sirve para la unión muscular y el almacenamiento de calcio. Algunos gasterópodos aparecen sin caparazón ( babosas ), pero pueden tener un remanente dentro del manto, o el caparazón se reduce de tal manera que el cuerpo no puede retraerse dentro ( semi-babosa ). Algunos caracoles también poseen un opérculo que sella la abertura de la concha, conocida como abertura , que brinda mayor protección. El estudio de las conchas de los moluscos se conoce como conchología . El estudio biológico de los gasterópodos, y de otros moluscos en general, es la malacología . Los términos de morfología de la concha varían según el grupo de especies.
Capas de concha
El caparazón de los gasterópodos tiene tres capas principales secretadas por el manto . La capa central calcárea, tracum, generalmente está hecha de carbonato de calcio precipitado en una matriz orgánica conocida como conquiolina . La capa más externa es el periostracum, que es resistente a la abrasión y proporciona la mayor parte de la coloración de la cáscara. El cuerpo del caracol entra en contacto con la capa lisa más interna que puede estar compuesta de madreperla o nácar de concha, una forma densa de conquiolina empaquetada horizontalmente, que se deposita sobre el periostraco a medida que crece el caracol.
Morfología
 |
|
|
| Morfología de la típica concha enrollada en espiral. El caparazón de Zonitoides nitidus , un caracol terrestre, tiene un enrollamiento dextral. Imagen superior: Vista dorsal, mostrando los verticilos y el ápice |
Foto de la concha de Zonitoides nitidus con una vista apical, una vista de apertura y una vista basal |
| Parte de una serie relacionada con |
| Biomineralización |
|---|
 |
La morfología de la concha de los gasterópodos suele ser bastante constante entre los individuos de una especie. Las variables de control son:
- La tasa de crecimiento por revolución alrededor del eje de bobinado. Las tasas altas dan formas de boca ancha como el abulón , las tasas bajas dan formas muy enrolladas como Turritella o algunas Planorbidae .
- La forma de la curva generadora, aproximadamente equivalente a la forma de la apertura. Puede ser redondo, por ejemplo en la concha turbante , alargado como en la concha cónica o tener una forma irregular con una extensión de canal sifonal, como en el Murex .
- La tasa de traducción de la curva de generación a lo largo del eje de enrollamiento, controlando qué tan alto se vuelve el caparazón resultante. Esto puede variar desde cero, una capa planispiral plana, hasta casi el diámetro de la apertura.
- Irregularidades o "esculturas" tales como costillas, espinas, protuberancias y várices hechas por el caracol cambiando regularmente la forma de la curva de generación durante el curso del crecimiento, por ejemplo en muchas especies de Murex .
- El crecimiento ontológico cambia a medida que el animal alcanza la edad adulta. Buenos ejemplos son el labio ensanchado del caracol adulto y el labio enrollado hacia adentro del cauri .
Algunos de estos factores se pueden modelar matemáticamente y existen programas para generar imágenes extremadamente realistas. Los primeros trabajos de David Raup en la computadora analógica también revelaron muchas combinaciones posibles que nunca fueron adoptadas por ningún gasterópodo real.
Algunas formas de concha se encuentran con mayor frecuencia en ciertos entornos, aunque hay muchas excepciones. Los ambientes de alta energía lavados por las olas, como la zona intermareal rocosa , suelen estar habitados por caracoles cuyas conchas tienen una amplia apertura, un área de superficie relativamente baja y una alta tasa de crecimiento por revolución. Las formas de agujas altas y muy esculpidas se vuelven más comunes en ambientes de aguas tranquilas. El caparazón de las formas excavadoras, como el olivo y la terebra , es liso, alargado y carece de una escultura elaborada, para disminuir la resistencia al moverse a través de la arena. En tierra, las formas de agujas altas a menudo se asocian con superficies verticales, mientras que los caracoles de caparazón plano tienden a vivir en el suelo.
Algunos gasterópodos, por ejemplo los Vermetidae , cementan la concha y crecen a lo largo de superficies sólidas como rocas u otras conchas.
quiralidad
La mayoría de las conchas de los gasterópodos están enrolladas en espiral. La mayoría (más del 90%) de las especies de gasterópodos tienen caparazones dextrales (diestros), pero una pequeña minoría de especies y géneros son prácticamente siempre sinistrales (zurdos), y muy pocas especies (por ejemplo , Amphidromus perversus ) muestran un caparazón dextral. mezcla de individuos dextrales y sinistrales. También se encuentran formas aberrantemente sinistrales de especies dextrales y algunas de ellas son muy buscadas por los recolectores de conchas.

Si se sostiene un caparazón de gasterópodo enrollado con la aguja apuntando hacia arriba y la abertura más o menos mirando hacia el observador, un caparazón dextral tendrá la abertura en el lado derecho y un caparazón sinistral tendrá la abertura en el lado izquierdo. . Esta quiralidad de los gasterópodos a veces se pasa por alto cuando las fotografías de gasterópodos enrollados son "volteadas" por un no experto antes de ser utilizadas en una publicación. Esta imagen "volteando" da como resultado que un gasterópodo dextral normal parezca ser uno sinistral raro o anormal.
La sinistralidad surgió de forma independiente 19 veces entre los gasterópodos marinos desde el inicio del Cenozoico . Esta zurdera parece ser más común en los pulmonados terrestres y de agua dulce. Pero aún así, las especies vivas dextrales en los gasterópodos parecen representar el 99% del número total.
La quiralidad en los gasterópodos aparece en la escisión temprana (escisión en espiral ) y el gen NODAL está involucrado. Un estudio más reciente (2013) correlaciona el enrollamiento asimétrico del caparazón con la expresión asimétrica izquierda-derecha del gen decapentapléjico en el manto.
Poblaciones mixtas en espiral
En unos pocos casos, se encuentran enrollamientos tanto para la mano izquierda como para la derecha en la misma población. Los mutantes sinistrales de especies normalmente dextrales y los mutantes dextrales de especies normalmente sinistrales son casos raros pero bien documentados entre los caracoles terrestres en general. Las poblaciones o especies con enrollamiento normalmente mixto son mucho más raras y, hasta donde se sabe, están confinadas, con una excepción, a unos pocos géneros de caracoles tropicales arbóreos. Además de Amphidromus , se conocen el Liguus vittatus cubano (Swainson), el Liguus virgineus haitiano (Linnaeus) (familia Orthalicidae ), algunas Partulina hawaianas y muchas Achatinella hawaianas (familia Achatinellidae ), así como varias especies de Partula de las islas del Pacífico (familia Partulidae ). tener poblaciones mixtas dextral-sinistral.
Una posible excepción puede referirse a algunos de los clausilidos europeos de la subfamilia Alopiinae . Son calcófilos obligados que viven en colonias aisladas sobre afloramientos calcáreos. Varios conjuntos de especies difieren solo en la dirección de enrollamiento, pero la evidencia no es concluyente en cuanto a si las conchas izquierdas y derechas viven juntas. Soos (1928, pp. 372–385) resumió las discusiones previas del problema y concluyó que las poblaciones de diestros y zurdos eran especies distintas. Otros han declarado que estas poblaciones no eran distintas y la cuestión está lejos de resolverse. El clausílido peruano, Nenia callistoglypta Pilsbry (1949, págs. 216–217), también se ha descrito como una especie de anfidromina.
Degner (1952) ha estudiado la genética del enrollamiento inverso en un raro mutante dextral de otro clausílido, Alinda biplicata (Montagu). El mecanismo es el mismo que en Radix peregra (Müller), con la dirección de enrollamiento determinada por un recesivo mendeliano simple.
Formas estándar de ver un caparazón

Vista de apertura de la concha de Valvata sincera

Vista panorámica de la concha de Valvata sincera

Vista umbilical de la concha de Valvata sincera

Esta vista dorsal del animal vivo Calliostoma bairdii también muestra una vista apical de su caparazón.




En fotografías o ilustraciones, el caparazón de un gasterópodo se puede mostrar orientado de varias maneras estándar:
- vista de apertura : este es el ángulo de visión más común. El caparazón se muestra en su totalidad, con su abertura hacia el espectador y el vértice en la parte superior. Si la abertura está en el lado derecho cuando se ve así, entonces el enrollamiento de la carcasa es "hacia la derecha" o dextral; si la abertura está en el lado izquierdo cuando se ve así, el caparazón tiene un enrollamiento de caparazón "zurdo" o sinistral.
- Vista apertural (o vista dorsal ): la concha se muestra con su apertura a 180° del observador y con el vértice en la parte superior.
- vista umbilical (o vista basal ): la concha se muestra vista directamente desde abajo. En la mayoría de los casos donde hay un ombligo, este está a la vista.
- vista apical : la concha se muestra mirando hacia abajo directamente sobre el ápice.
Descripción

El caparazón comienza con el caparazón de la larva, los verticilos embrionarios (generalmente) diminutos conocidos como protoconcha , que a menudo es bastante distinto del resto del caparazón y no tiene líneas de crecimiento. A partir de la protoconcha, que forma el vértice de la aguja , las espirales o verticilos de la concha aumentan gradualmente de tamaño. Normalmente, los verticilos son de sección circular o elíptica. La aguja puede ser alta o baja, ancha o esbelta, según la forma en que estén dispuestas las espirales de la concha, y el ángulo apical de la concha varía en consecuencia. Los verticilos a veces descansan libremente unos sobre otros (como en Epitonium scalare ). También pueden superponerse a los verticilos anteriores, de modo que los verticilos anteriores pueden quedar cubiertos en gran parte o en su totalidad por los posteriores. Cuando se produce una angulación, el espacio entre ésta y la sutura situada encima constituye el área conocida como "hombro" de la cubierta. El ángulo del hombro puede ser liso o en quilla y, a veces, puede tener nudos o espinas.
La forma más simple de escultura del caparazón de gasterópodo consiste en crestas longitudinales y/o crestas transversales. Las espirales primarias pueden aparecer en sucesión regular a ambos lados de la primera primaria, que generalmente se convierte en el ángulo del hombro si se produce una angulación. Las espirales secundarias pueden aparecer por intercalación entre las primarias y generalmente están ausentes en la concha joven, excepto en algunos tipos muy acelerados. Las espirales terciarias se intercalan entre los grupos precedentes en especies más especializadas. Las costillas son pliegues transversales regulares de la cubierta, que generalmente se extienden de sutura a sutura. Suelen estar espaciados uniformemente y atravesados por las espirales. En tipos especializados, cuando se forma un ángulo del hombro, se concentran como nodos sobre este ángulo, desapareciendo del hombro arriba y del cuerpo abajo. Las espinas pueden reemplazar los nodos en etapas posteriores. Se forman como muescas en el margen de la concha y posteriormente se abandonan, quedando a menudo abiertas por delante. También pueden surgir espinas irregulares en varias partes de la superficie del caparazón (ver Platyceras ).
Cuando se forma una fila de espinas en el borde o en el borde exterior de la concha durante un período de descanso, esta característica a veces permanece como una várice como en ( Murex ) y muchos de los Ranellidae . Las várices también pueden formarse por la simple expansión del labio externo y una posterior reanudación del crecimiento desde la base de la expansión.
La abertura o peristoma de la cubierta puede ser simple o modificarse de diversas formas. Generalmente se reconocen un labio externo y uno interno (columelar). Estos pueden ser continuos entre sí o pueden estar divididos por una muesca anterior. Este, en algunos tipos ( Fusinus , etc.) se extrae hacia un canal sifonal anterior , de mayor o menor longitud.
Una muesca superior o posterior está presente en ciertos taxones, y esto puede resultar en la formación de una cresta o plataforma al lado de la sutura ( Cavilithes ). En ciertos tipos (Pleurotomidae, Pleurotomaridae, Bellerophontidae, etc.), se produce una emarginación o muesca externa (lateral), a veces prolongada en una hendidura, y el cierre progresivo de esta hendidura puede dar lugar a una banda de hendidura claramente marcada. En algunos casos la hendidura es abandonada y dejada como un agujero ( Fissurellidae ), o por renovación periódica como una sucesión de agujeros ( Haliotis ). La emarginación externa a menudo solo se indica por el curso reflejado de las líneas de crecimiento en el caparazón.
En el interior del labio externo, a veces se encuentran varias crestas o pliegues llamados liras , y estos ocasionalmente pueden ser fuertes y con forma de dientes ( Nerinea ). Crestas o pliegues o pliegues columelares similares se encuentran con mayor frecuencia en el labio interno, junto a la columela o giro espiral central. Estos pueden ser oblicuos o normales al eje de enrollamiento (horizontal), pocos o numerosos, fácilmente visibles, o lejos dentro de la concha para ser invisibles excepto en conchas rotas. Cuando el eje de enrollamiento es hueco (aguja perforada), la abertura en la base constituye el ombligo . El ombligo varía mucho en tamaño y puede estar total o parcialmente cubierto por una expansión o callo del labio interno ( Natica ).
Muchos caparazones recientes , cuando el animal está vivo o el caparazón está recién vacío, tienen una capa superior de epidermis o periostraco córnea, suave o peluda , una capa proteica que a veces es lo suficientemente gruesa como para ocultar las marcas de color de la superficie del caparazón. cascarón. El periostracum, así como la coloración, rara vez se conservan en conchas fósiles.
El extremo de apertura del caparazón de los gasterópodos es el extremo anterior, el más cercano a la cabeza del animal; el vértice de la aguja es a menudo el extremo posterior o al menos es el lado dorsal. La mayoría de los autores calculan las conchas con el vértice de la aguja hacia arriba. En vida, cuando las partes blandas de estos caracoles se retraen, en algunos grupos la abertura de la concha se cierra mediante el uso de un opérculo córneo o calcáreo , una estructura similar a una puerta secretada por la superficie superior del caracol y adherida a ella. parte posterior del pie. El opérculo es de forma muy variable en los diferentes grupos de caracoles que lo poseen.
Partes de la concha

La terminología utilizada para describir las conchas de los gasterópodos incluye:
- Apertura : la apertura de la concha
- Labio : peristoma : el margen de la apertura
- Ápice : los pocos verticilos más pequeños de la concha.
- Verticilo corporal (o último verticilo): el verticilo de mayor tamaño en el que se encuentra la mayor parte de la masa visceral del molusco.
- Columella : la "pequeña columna" en el eje de revolución de la concha
- Opérculo : la "trampilla" de la concha
- Callo parietal : una cresta en el labio interior de la abertura en ciertos gasterópodos
- Periostracum : una capa delgada de "piel" orgánica que forma la capa exterior del caparazón de muchas especies.
- Peristoma : la parte del caparazón que se encuentra justo alrededor de la abertura, también conocida como labio.
- Trenza : pliegues en la columela.
- Protoconch : el verticilo nuclear o embrionario; el caparazón de la larva, a menudo permanece en posición incluso en un caparazón adulto
-
Escultura : ornamentación en la superficie exterior de una concha.
- Lira : líneas elevadas o crestas en la superficie de la concha
- Canal sifonal : una extensión de la abertura en ciertos gasterópodos.
- Spire : la parte de la concha sobre el verticilo del cuerpo.
- Sutura : la unión entre los verticilos de la mayoría de los gasterópodos .
- Teleoconch: toda la concha sin la protoconch; los verticilos posnucleares.
- Ombligo : en las conchas donde los verticilos se separan a medida que crecen, en la parte inferior de la concha hay una depresión profunda que llega hasta la aguja; este es el ombligo
- Varix : en algunas conchas de moluscos, costillas verticales elevadas y engrosadas espaciadas marcan el final de un período de rápido crecimiento; estas son las várices
- Espiral : cada una de las rotaciones completas de la espiral de la concha
Forma de la concha
La forma general de la concha varía. Por ejemplo, se pueden distinguir tres grupos en función de la relación alto-ancho:
- oblongo: la altura es mucho mayor que el ancho
- concha globosa o cónica: la altura y el ancho de la concha son aproximadamente iguales
- deprimido - el ancho es mucho mayor que la altura

concha oblonga de Bulgarica denticulata

concha globosa de Sphincterochila candidissima

concha deprimida de Escargot de Quimper


Las siguientes son las principales modificaciones de forma en el caparazón de los gasterópodos.
- Regularmente espiral:
- Bulloide: Bulla en forma de burbuja
- Coeloconoid un caparazón cónico ligeramente cóncavo en el que el ángulo incremental aumenta constantemente durante el crecimiento (ver: Calliostoma )
- En forma de cono, obcónico. Cono
- Contabular, corto, con verticilos a hombros
- Convoluta: abertura tan larga como la concha, que oculta casi o completamente la aguja. Cipraea
- Cilíndrica, pupiiforme. Lioplax , Pupa
- Cyrtoconoid: acercándose a una forma cónica pero con lados convexos (ver: Gibbula )
- Deprimido, lenticular. Ethalia carneolata
- Discoidal. Elacorbis
- En forma de oreja. Haliotis
- Alargado, subulado, elevado. Terebra
- Pocos verticilos. Hélix pomatia .
- Fusiforme, en forma de huso. Fusino
- Giboso. Los verticilos se hincharon más allá del contorno normal de aumento (generalmente en el lado de la apertura). estreptaxis .
- Globular. Natica
- Muchos verticilos. Millerelix peregrina .
- Corto, bucciniforme. buccinum
- Troquiforme, piramidal, cónica con base plana. troco
- Turbinado: cónico, con base redondeada. Turbo
- Torreón, turbulento, babilónico; una concha alargada con los verticilos angulados o con hombros en su parte superior. Turritella
- Escalariforme, verticilos que no chocan. epitonio escalar
- Irregularmente espiral, evoluciona. Siliquaria , Vermetus
- Tubular.
- En forma de escudo. umbráculo
- En forma de barco, en forma de zapatilla. Crepídula
- Cónico o en forma de lapa. Rótula
- Bicónico: en forma de dos formas cónicas que se tocan en sus bases y se estrechan en ambos extremos: Fasciolaria tulipa
- En forma de pera: combinación de dos formas: ovado-cónica y cónica. ficus
La distinción detallada de la forma puede ser:

forma de gorra

forma de oreja

neritiforme

planispiral

troquiforme deprimida o valvatiforme

troquiforme

ovado-cónico

cónico

forma alargada-cónica o turriforme o en espiral

plena forma

forma de huso: el caracol de mar Syrinx aruanus tiene el caparazón más grande de todos los gasterópodos vivos.

forma de maza - Venus Peine Murex

forma de huevo

Forma irregular














Dimensiones
Las medidas más utilizadas de la concha de un gasterópodo son: la altura de la concha, la anchura de la concha, la altura de la abertura y la anchura de la abertura. El número de verticilos también se usa a menudo.
En este contexto, la altura (o la longitud) de una concha es su medida máxima a lo largo del eje central. El ancho (o ancho, o diámetro) es la medida máxima de la concha en ángulo recto con el eje central. Ambos términos solo están relacionados con la descripción del caparazón y no con la orientación del caparazón en el animal vivo.
La mayor altura de cualquier caparazón se encuentra en la especie de caracol marino Syrinx aruanus , que puede alcanzar los 91 cm.
El eje central es un eje imaginario a lo largo de una concha, alrededor del cual, en una concha enrollada, giran los verticilos. El eje central pasa por la columela, el pilar central de la concha.
Cambios evolutivos
Entre los roles propuestos invocados para la variabilidad de las conchas durante la evolución se incluyen la estabilidad mecánica, la defensa contra los depredadores y la selección climática.
Las conchas de algunos gasterópodos se han reducido total o parcialmente durante su evolución . Esta reducción se puede ver en todas las babosas , en las semibabosas y en varios otros gasterópodos marinos y no marinos. En ocasiones, la reducción del caparazón se asocia con una forma de alimentación depredadora.
Algunos taxones perdieron el enrollamiento de su caparazón durante la evolución. De acuerdo con la ley de Dollo , no es posible recuperar el enrollamiento del cascarón después de que se haya perdido. A pesar de eso, hay pocos géneros en la familia Calyptraeidae que cambiaron su tiempo de desarrollo ( heterocronía ) y recuperaron ( reevolución ) un caparazón enrollado desde la condición anterior de un caparazón desenrollado similar a una lapa .
Implicaciones tafonómicas
En cantidades suficientemente grandes, las conchas de gasterópodos pueden tener un impacto suficiente en las condiciones ambientales para afectar la capacidad de fosilizar de los restos orgánicos en el entorno local. Por ejemplo, en la Formación Dinosaur Park , la cáscara de huevo fósil de hadrosaurio es rara. Esto se debe a que la descomposición de los taninos de la vegetación local de coníferas habría causado que las antiguas aguas se volvieran ácidas. Los fragmentos de cáscara de huevo están presentes en solo dos sitios de microfósiles , en los cuales predominan las cáscaras conservadas de vida de invertebrados, incluidos los gasterópodos. Fue la lenta disolución de estas cáscaras liberando carbonato de calcio en el agua lo que elevó el pH del agua lo suficientemente alto como para evitar que los fragmentos de cáscara de huevo se disolvieran antes de que pudieran fosilizarse.
Variedad de formas

Turritella communis , concha de muchos verticilos de caracol torre

Imagen de rayos X de Turritella

Concha de caracol cauri marino - Cypraea nebrites

Concha espiral de epitonio escalar





Imagen de rayos X de Cypraea

Imagen de rayos X de la concha de Tonna galea
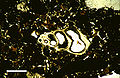
Sección delgada en luz polarizada plana de caparazón de gasterópodo microscópico, del sedimento lagunar del Holoceno de Rice Bay, isla de San Salvador , Bahamas. Barra de escala 500 µm.





Referencias
Este artículo incorpora texto de dominio público de las referencias y texto CC-BY-2.0 de la referencia.
Otras lecturas
| Parte de una serie sobre |
| conchas marinas |
|---|
 |
| conchas de moluscos |
| Sobre las conchas de moluscos |
| Otras conchas marinas |
- Acerca de la quiralidad
- van Batenburg1 FHD y Gittenberger E. (1996). "Facilidad de fijación de un cambio en el enrollamiento: experimentos informáticos sobre quiralidad en caracoles". Herencia 76 : 278–286. doi : 10.1038/hdy.1996.41 .
- Wandelt J. & Nagy LM (24 de agosto de 2004) "Asimetría izquierda-derecha: más de una forma de enrollar una concha". Biología actual 14 (16): R654–R656. doi : 10.1016/j.cub.2004.08.010
enlaces externos
![]() Medios relacionados con las conchas de Gasterópodos en Wikimedia Commons
Medios relacionados con las conchas de Gasterópodos en Wikimedia Commons
- Gasterópodos por JH Leal – Información sobre algunos gasterópodos del Atlántico occidental tropical, específicamente el Mar Caribe, con relevancia para las pesquerías en esa región
- Datación por radiocarbono de conchas de gasterópodos
- Nair KK & Muthe PT (18 de noviembre de 1961) "Efecto de la ribonucleasa en la regeneración de la concha en Ariophanta sp." . Naturaleza 192 : 674–675. doi : 10.1038/192674b0 .
- (en español) Antonio Ruiz Ruiz, Ángel Cárcaba Pozo, Ana I. Porras Crevillen & José R. Arrébola Burgos Caracoles Terrestres de Andalúcia. Guia y manual de identificacion . 303 págs., ISBN 84-935194-2-1 . ( del sitio web )