
Anatomía del cerebelo - Anatomy of the cerebellum
| Cerebelo | |
|---|---|
 Dibujo del cerebro humano, que muestra el cerebelo y la protuberancia
| |

Corte transversal vertical de la línea media del cerebelo humano, que muestra el patrón de plegado de la corteza y las estructuras interiores
| |
| Detalles | |
| Parte de | Metencéfalo |
| Artería | SCA , AICA , PICA |
| Vena | superior , inferior |
| Identificadores | |
| Identificación de NeuroLex | birnlex_1489 |
| TA98 | A14.1.07.001 |
| TA2 | 5788 |
| Términos anatómicos de la neuroanatomía | |
La anatomía del cerebelo se puede ver en tres niveles. En el nivel de la anatomía macroscópica , el cerebelo consta de una capa de corteza arrugada y muy doblada, con sustancia blanca debajo, varios núcleos profundos incrustados en la sustancia blanca y un ventrículo lleno de líquido en el medio. En el nivel intermedio, el cerebelo y sus estructuras auxiliares se pueden dividir en varios cientos o miles de módulos o compartimentos que funcionan de forma independiente, conocidos como microzonas . A nivel microscópico, cada módulo consta del mismo pequeño conjunto de elementos neuronales, dispuestos con una geometría altamente estereotipada.
Anatomia asquerosa
El cerebelo está ubicado en la base del cerebro, con la gran masa de la corteza cerebral por encima y la porción del tronco del encéfalo llamada protuberancia frente a él. Está separada del cerebro suprayacente por una capa de duramadre resistente ; todas sus conexiones con otras partes del cerebro viajan a través de la protuberancia. Los anatomistas clasifican el cerebelo como parte del metencéfalo , que también incluye la protuberancia; el metencéfalo, a su vez, es la parte superior del rombencéfalo o "rombencéfalo". Como la corteza cerebral, el cerebelo se divide en dos hemisferios; también contiene una zona estrecha en la línea media llamada vermis . Se usa convencionalmente un conjunto de pliegues grandes para dividir la estructura general en diez lóbulos más pequeños .
Debido a su gran cantidad de células granulares diminutas , el cerebelo contiene más neuronas que el resto del cerebro en su conjunto, pero solo ocupa el 10% del volumen total del cerebro. El cerebelo recibe cerca de 200 millones de fibras de entrada; por el contrario, el nervio óptico está compuesto por apenas un millón de fibras.
El aspecto inusual de la superficie del cerebelo oculta el hecho de que la mayor parte de la estructura está formada por una capa muy apretada de materia gris, la corteza cerebelosa . Se ha estimado que si la corteza cerebelosa humana pudiera desplegarse por completo, daría lugar a una capa de tejido neural de aproximadamente 1 metro de largo y 10 centímetros de ancho, una superficie total de 500-1000 cm cuadrados, todo empaquetado dentro de un volumen de 100-150 cm cúbicos Debajo de la sustancia gris de la corteza se encuentra la sustancia blanca , formada en gran parte por fibras nerviosas mielinizadas que van hacia y desde la corteza. Incrustados dentro de la sustancia blanca, que a veces se llama arbor vitae (árbol de la vida) en el cerebelo debido a su apariencia ramificada, hay cuatro núcleos cerebelosos profundos .
El cerebelo se puede dividir de acuerdo con tres criterios diferentes: anatómico macroscópico, filogenético y funcional.
Divisiones anatómicas macroscópicas
A simple vista, se pueden distinguir tres lóbulos en el cerebelo: el lóbulo floculonodular , el lóbulo anterior (rostral a la "fisura primaria") y el lóbulo posterior (dorsal a la "fisura primaria"). Los dos últimos se pueden dividir en un vermis cerebeloso de línea media y hemisferios cerebelosos laterales.

Figura 3: Cerebelo y regiones circundantes; vista sagital de un hemisferio. A: mesencéfalo . B: Pons . C: médula . D: médula espinal . E: cuarto ventrículo . F: Arbor vitae . G: Amígdalas . H: lóbulo anterior. I: lóbulo posterior.
|

Divisiones filogenéticas y funcionales
El cerebelo también se puede dividir en tres partes basándose tanto en criterios filogenéticos (la edad evolutiva de cada parte) como en criterios funcionales (las conexiones entrantes y salientes que tiene cada parte y el papel que desempeña en la función cerebelosa normal). Desde el más antiguo filogenéticamente al más nuevo, las tres partes son:
| Denominación funcional ( denominación filogenética) | Partes anatómicas | Papel |
| Vestibulocerebellum (Archicerebellum) | Lóbulo floculonodular (y vermis inmediatamente adyacente) | El vestibulocerebelo regula el equilibrio y los movimientos oculares. Recibe información vestibular tanto de los canales semicirculares como de los núcleos vestibulares , y envía fibras de regreso a los núcleos vestibulares medial y lateral. También recibe información visual de los colículos superiores y de la corteza visual (esta última a través de los núcleos pontinos , formando una vía cortico-ponto-cerebelosa). Las lesiones del vestibulocerebelo provocan alteraciones del equilibrio y la marcha . Hay otra pequeña región, conocida como lóbulo biventer . |
| Espinocerebelo (paleocerebelo) | Vermis y partes intermedias de los hemisferios ("paravermis") | El espinocerebelo regula los movimientos del cuerpo y las extremidades. Recibe información de la propiocepción de las columnas dorsales de la médula espinal (incluido el tracto espinocerebeloso ) y del nervio trigémino , así como de los sistemas visual y auditivo . Envía fibras a los núcleos cerebelosos profundos (incluido el núcleo fastigial ) que a su vez se proyectan tanto a la corteza cerebral (a través del mesencéfalo y el tálamo ) como al tallo cerebral (a través de la formación reticular en la protuberancia y los núcleos vestibulares en el bulbo raquídeo ), por lo tanto proporcionando modulación de sistemas de motores descendentes. El espinocerebelo contiene mapas sensoriales, ya que recibe datos sobre la posición de varias partes del cuerpo en el espacio: en particular, el vermis recibe fibras del tronco y las porciones proximales de las extremidades, mientras que las partes intermedias de los hemisferios reciben fibras de las porciones distales de las extremidades. . El espinocerebelo es capaz de elaborar información propioceptiva para anticipar la posición futura de una parte del cuerpo durante el curso de un movimiento, de una manera "retroalimentada". |
| Cerebrocerebelo (Neocerebelo, Pontocerebelo) | Partes laterales de los hemisferios | El neocerebelo participa en la planificación del movimiento y la evaluación de la información sensorial para la acción. Recibe información exclusivamente de la corteza cerebral (especialmente el lóbulo parietal ) a través de los núcleos pontinos (en la protuberancia , formando vías cortico-ponto-cerebelosas) y el núcleo dentado (en el cerebelo ), y envía fibras principalmente al tálamo ventrolateral (en a su vez conectado a las áreas motoras de la corteza premotora y al área motora primaria de la corteza cerebral) y al núcleo rojo (a su vez conectado al núcleo olivar inferior , que se une de nuevo a los hemisferios cerebelosos). El neocerebelo participa en la planificación del movimiento que está a punto de ocurrir y también tiene funciones puramente cognitivas. |
Mucho de lo que se entiende sobre las funciones del cerebelo se deriva de la documentación cuidadosa de los efectos de las lesiones focales en pacientes humanos que han sufrido una lesión o enfermedad o mediante la investigación de lesiones en animales.
Anatomía celular
Como se explica con más detalle en la sección Función , el cerebelo se diferencia de la mayoría de las otras áreas del cerebro en que el flujo de señales neuronales a través de él es casi completamente unidireccional: prácticamente no existen conexiones hacia atrás entre sus elementos neuronales. Por lo tanto, la forma más lógica de describir la estructura celular es comenzar con las entradas y seguir la secuencia de conexiones hasta las salidas.
Núcleos profundos
Los cuatro núcleos profundos del cerebelo son los núcleos dentado , emboliforme , globoso y fastigii y actúan como los principales centros de comunicación, enviando y recibiendo información hacia y desde partes específicas del cerebro. Además, estos núcleos reciben señales inhibidoras y excitadoras de otras partes del cerebro que a su vez afectan las señales salientes del núcleo (los núcleos globoso y emboliforme forman el núcleo interpuesto ).
Capas corticales


La citoarquitectura ( organización celular ) del cerebelo es muy uniforme, con conexiones organizadas en una matriz tridimensional aproximada de elementos de circuito perpendiculares . Esta uniformidad organizativa hace que los circuitos nerviosos sean relativamente fáciles de estudiar.
Hay tres capas en la corteza cerebelosa; de la capa externa a la interna, estas son las capas molecular, de Purkinje y granular. La función de la corteza cerebelosa es esencialmente modular la información que fluye a través de los núcleos profundos. El microcircuito del cerebelo se esquematiza en la Figura 5. Las fibras musgosas y trepadoras transportan información sensoriomotora hacia los núcleos profundos, que a su vez la transmiten a varias áreas premotoras, regulando así la ganancia y la sincronización de las acciones motoras. Las fibras musgosas y trepadoras también alimentan esta información en la corteza cerebelosa, que realiza varios cálculos, lo que resulta en la regulación de la activación de las células de Purkinje. Las neuronas de Purkinje retroalimentan a los núcleos profundos a través de una potente sinapsis inhibidora . Esta sinapsis regula la medida en que las fibras musgosas y trepadoras activan los núcleos profundos y, por lo tanto, controlan el efecto final del cerebelo sobre la función motora. Se ha demostrado que la fuerza sináptica de casi todas las sinapsis de la corteza cerebelosa experimenta una plasticidad sináptica . Esto permite que los circuitos de la corteza cerebelosa ajusten y afinen continuamente la producción del cerebelo, formando la base de algunos tipos de aprendizaje motor y coordinación. Cada capa de la corteza cerebelosa contiene los diversos tipos de células que componen este circuito.
Capa molecular
Esta capa más externa de la corteza cerebelosa contiene dos tipos de interneuronas inhibidoras : las células estrelladas y en cesto . También contiene los cenadores dendríticos de las neuronas de Purkinje y tractos de fibra paralelos de las células granulares. Tanto las células estrelladas como las cestas forman sinapsis GABAérgicas en las dendritas de las células de Purkinje.
Capa de Purkinje
La capa intermedia contiene solo un tipo de cuerpo celular: el de la gran célula de Purkinje . Las células de Purkinje son las neuronas integradoras primarias de la corteza cerebelosa y proporcionan su única salida. Las dendritas de las células de Purkinje son grandes pérgolas con cientos de ramas espinosas que llegan hasta la capa molecular (Fig. 6). Estos cenadores dendríticos son planos (casi todos se encuentran en planos) con los cenadores vecinos de Purkinje en planos paralelos. Cada fibra paralela de las células granulares corre ortogonalmente a través de estos pilares, como un cable que atraviesa muchas capas. Las neuronas de Purkinje son GABAérgicas, lo que significa que tienen sinapsis inhibidoras, con las neuronas de los núcleos cerebelosos y vestibulares profundos del tronco del encéfalo. Cada celda de Purkinje recibe una entrada excitadora de 100.000 a 200.000 fibras paralelas. Se dice que las fibras paralelas son responsables de los picos simples (todo o nada, invariante de amplitud ) de la célula de Purkinje.
Las células de Purkinje también reciben información del núcleo olivar inferior a través de fibras trepadoras . Un buen mnemónico para esta interacción es la frase "trepa al otro olivo", dado que las fibras trepadoras se originan en la aceituna inferior contralateral. En sorprendente contraste con las más de 100.000 entradas de fibras paralelas, cada celda de Purkinje recibe información de exactamente una fibra trepadora; pero esta única fibra "trepa" por las dendritas de la célula de Purkinje, se enrolla alrededor de ellas y crea un gran número de sinapsis a medida que avanza. La entrada neta es tan fuerte que un solo potencial de acción de una fibra trepadora es capaz de producir un "pico complejo" en la celda de Purkinje: un estallido de varios picos seguidos, con amplitud decreciente, seguido de una pausa durante la cual picos simples están suprimidos.
Justo debajo de la capa de Purkinje se encuentran las células de Lugaro cuyas dendritas muy largas viajan a lo largo del límite entre Purkinje y las capas granulares.
Capa granular
La capa más interna contiene los cuerpos celulares de tres tipos de células: las numerosas y diminutas células granulares , las células en cepillo unipolares un poco más grandes y las células de Golgi, mucho más grandes . Las fibras musgosas entran en la capa granular desde su principal punto de origen, los núcleos pontinos. Estas fibras forman sinapsis excitadoras con las células granulares y las células de los núcleos cerebelosos profundos. Las células granulares envían sus axones en forma de T, conocidos como fibras paralelas, hacia la capa molecular superficial, donde forman cientos de miles de sinapsis con las dendritas de las células de Purkinje . El cerebelo humano contiene del orden de 60 a 80 mil millones de células granulosas, lo que hace que este tipo de célula única sea , con mucho, la neurona más numerosa del cerebro (aproximadamente el 70% de todas las neuronas del cerebro y la médula espinal, combinadas). Las células de Golgi proporcionan retroalimentación inhibidora a las células granulares, formando una sinapsis con ellas y proyectando un axón hacia la capa molecular.
Relación con la corteza cerebral
Los potenciales de campo local de la neocorteza y el cerebelo oscilan coherentemente a (6–40 Hz) en animales que se comportan despiertos. Estos parecen estar bajo el control de la producción de la corteza cerebral. Esta salida estaría mediada por una vía desde la capa 5/6 de neuronas en la neocorteza a través de ese proyecto hasta la protuberancia o la aceituna inferior. Si a través de la protuberancia esto iría a las fibras musgosas que hacen sinapsis con las neuronas granulares y de Golgi con las células granulares, entonces se dirigen a las neuronas de Purkinje a través de sus fibras paralelas excitadoras. Si la aceituna inferior iría a través de entradas de fibra trepadora excitadoras a las neuronas de Purkinje. Estos devuelven esta salida a la corteza cerebral a través del tálamo ventrolateral completando el bucle.
La vía corticopontocerebelosa es la vía más grande asociada con el cerebelo. Surgiendo en la corteza cerebral, estas fibras primero terminan ipsilateralmente en los núcleos pontinos . Luego, las fibras se decusan y forman el pedúnculo cerebeloso medio, terminando en la corteza cerebelosa como fibras musgosas. Esta vía transmite señales que informan al cerebelo sobre el movimiento en curso y el próximo movimiento. Esto ayuda al ajuste continuo de la actividad motora.
El inicio del movimiento se transmite al cerebelo a través de la vía corticoreticulocerebelosa. Estos sinapsis ipsilateralmente en la formación reticular , luego a través de los pedúnculos inferior y medio hacia el vermis cerebeloso .
La corteza motora y la corteza somatosensorial se proyecta sobre los núcleos olivar inferior y accesorio ipsilaterales, formando luego el tracto olivocerebeloso . Las fibras cortico-olivares hacen sinapsis bilateralmente en el núcleo olivar inferior . El orden se conserva en las proyecciones del tracto olivocerebeloso sobre los "mapas corporales" de la corteza cerebelosa contralateral. En condiciones de reposo en experimentos con animales, grupos de neuronas olivares se descargan sincrónicamente a 5 a 10 Hz (impulsos / s). En la corteza cerebelosa, la respuesta de las células de Purkinje toma la forma de picos complejos.
El cerebelo envía sus proyecciones de regreso a la corteza cerebral a través del tracto cerebelotalámico .
La expansión lateral cerebelosa, o neocerebelo, puede estar asociada con funciones cognitivas y está anatómicamente vinculada con la corteza prefrontal lateral . Muestra mayor actividad durante el habla, con predominio unilateral consistente con una posible vinculación (a través del tálamo) con el área motora del habla.
Cuando ocurren lesiones en las áreas de asociación ligadas al cerebelo por fibras corticopontocerebelosas, puede ocurrir el síndrome cognitivo afectivo. Esto da como resultado defectos cognitivos en forma de poder de razonamiento disminuido, falta de atención, errores gramaticales en el habla, sentido espacial deficiente y pérdida de memoria irregular.
Suministro de sangre


Tres arterias suministran sangre al cerebelo (Fig. 7): la arteria cerebelosa superior (SCA), la arteria cerebelosa anteroinferior (AICA) y la arteria cerebelosa posteroinferior (PICA).
El SCA se ramifica desde la porción lateral de la arteria basilar, justo por debajo de su bifurcación hacia la arteria cerebral posterior . Aquí, se envuelve posteriormente alrededor de la protuberancia (a la que también suministra sangre) antes de llegar al cerebelo. El SCA suministra sangre a la mayor parte de la corteza cerebelosa, los núcleos cerebelosos y los pedúnculos cerebelosos superiores.
La AICA se ramifica desde la porción lateral de la arteria basilar, justo por encima de la unión de las arterias vertebrales. Desde su origen, se ramifica a lo largo de la porción inferior de la protuberancia en el ángulo pontocerebeloso antes de llegar al cerebelo. Esta arteria irriga la porción anterior del cerebelo inferior, el pedúnculo cerebeloso medio y los nervios facial (VII par) y vestibulococlear (VIII par). La obstrucción del AICA puede causar paresia , parálisis y pérdida de sensibilidad en la cara; también puede causar problemas de audición . Además, podría provocar un infarto del ángulo pontocerebeloso. Esto podría dar lugar a hiperacusia (disfunción del músculo estapedio , inervado por el par VII ) y vértigo (interpretación incorrecta de la aceleración de la endolinfa del canal semicircular vestibular causada por la alteración del VIII par ).
La PICA se ramifica desde la porción lateral de las arterias vertebrales justo por debajo de su unión con la arteria basilar. Antes de llegar a la superficie inferior del cerebelo, el PICA envía ramas hacia la médula, suministrando sangre a varios núcleos de pares craneales . En el cerebelo, el PICA suministra sangre a la porción posteroinferior del cerebelo, el pedúnculo cerebeloso inferior, el núcleo ambiguo , el núcleo motor vago, el núcleo espinal del trigémino , el núcleo solitario y los núcleos vestibulococleares .
Variaciones entre vertebrados
Existe una variación considerable en el tamaño y la forma del cerebelo en diferentes especies de vertebrados. Generalmente es más grande en peces , aves y mamíferos cartilaginosos y óseos , pero algo más pequeño en reptiles. Los lóbulos grandes emparejados y contorneados que se encuentran en los seres humanos son típicos de los mamíferos, pero el cerebelo es generalmente un lóbulo mediano único en otros grupos, y es liso o solo ligeramente acanalado. En los mamíferos, el neocerebelo es la mayor parte del cerebelo en masa, pero en otros vertebrados suele ser el espinocerebelo.
En anfibios , lampreas y brujas, el cerebelo está poco desarrollado; en los dos últimos grupos apenas se distingue del tronco encefálico. Aunque el espinocerebelo está presente en estos grupos, las estructuras primarias son pequeños núcleos pareados correspondientes al vestibulocerebelo.
Pedúnculos
El cerebelo sigue el patrón general de grupos de tres que se encuentra en la anatomía, con tres pedúnculos cerebelosos principales de entrada y salida (haces de fibras). Estos son los pedúnculos cerebelosos superior (brachium conjunctivum), medio (brachium pontis) e inferior ( cuerpos restiformes y yuxtarestiformes ).
| Pedúnculo | Descripción |
| Superior | Si bien hay algunas fibras aferentes del tracto espinocerebeloso anterior que se transportan al lóbulo cerebeloso anterior a través de este pedúnculo, la mayoría de las fibras son eferentes. Por tanto, el pedúnculo cerebeloso superior es la principal vía de salida del cerebelo. La mayoría de las fibras eferentes se originan dentro del núcleo dentado que, a su vez, se proyecta a varias estructuras del mesencéfalo , incluido el núcleo rojo , el núcleo ventral lateral / ventral anterior del tálamo y la médula . Las vías dentatorubrothalamocortical (núcleo dentado> núcleo rojo > tálamo > corteza premotora ) y cerebelotalamocortical (cerebelo> tálamo> corteza premotora) son dos vías principales que atraviesan este pedúnculo y son importantes en la planificación motora . |
| Medio | Está compuesto en su totalidad por fibras aferentes que se originan dentro de los núcleos pontinos como parte del tracto corticopontocerebeloso masivo (corteza cerebral> protuberancia> cerebelo). Estas fibras descienden de las áreas sensoriales y motoras del neocórtex cerebral y hacen que el pedúnculo cerebeloso medio sea el más grande de los tres pedúnculos cerebelosos. |
| Inferior | Este transporta muchos tipos de fibras de entrada y salida que se ocupan principalmente de integrar la entrada sensorial propioceptiva con funciones vestibulares motoras como el equilibrio y el mantenimiento de la postura. La información propioceptiva del cuerpo se lleva al cerebelo a través del tracto espinocerebeloso dorsal . Este tracto pasa a través del pedúnculo cerebeloso inferior y hace sinapsis dentro del paleocerebelo. La información vestibular se proyecta sobre el archicerebelo. Las fibras trepadoras de la aceituna inferior corren a través del pedúnculo cerebeloso inferior. Este pedúnculo también transporta información directamente desde las células de Purkinje a los núcleos vestibulares en el tallo cerebral dorsal ubicado en la unión entre la protuberancia y la médula . |
Hay tres fuentes de entrada al cerebelo, en dos categorías que consisten en fibras musgosas y trepadoras, respectivamente. Las fibras musgosas pueden originarse en los núcleos pontinos, que son grupos de neuronas ubicadas en la protuberancia que transportan información de la corteza cerebral contralateral. También pueden surgir dentro del haz espinocerebeloso cuyo origen se localiza en la médula espinal ipsolateral . La mayor parte de la salida del cerebelo inicialmente hace sinapsis en los núcleos cerebelosos profundos antes de salir a través de los tres pedúnculos. La excepción más notable es la inhibición directa de los núcleos vestibulares por las células de Purkinje.
Desarrollo
Durante las primeras etapas del desarrollo embrionario , el cerebro comienza a formarse en tres segmentos distintos: prosencéfalo , mesencéfalo y rombencéfalo . El rombencéfalo es el segmento más caudal (hacia la cola) del cerebro embrionario; es a partir de este segmento que se desarrolla el cerebelo. A lo largo del segmento embrionario rombencefálico se desarrollan ocho inflamaciones, llamadas rombómeros . El cerebelo surge de dos rombómeros ubicados en la placa alar del tubo neural , una estructura que eventualmente forma el cerebro y la médula espinal. Los rombómeros específicos a partir de los cuales se forma el cerebelo son el rombómero 1 (Rh.1) caudalmente (cerca de la cola) y el "istmo" rostral (cerca del frente).
Se cree que dos regiones primarias dan lugar a las neuronas que forman el cerebelo. La primera región es la zona ventricular en el techo del cuarto ventrículo . Esta área produce células de Purkinje y neuronas nucleares cerebelosas profundas . Estas células son las neuronas de salida primarias de la corteza cerebelosa y el cerebelo. La segunda zona germinal (lugar de nacimiento celular) se conoce como el labio rómbico, luego las neuronas se mueven en la semana 27 del embrión humano hacia la capa granular externa . Esta capa de células, que se encuentra en el exterior del cerebelo, produce las neuronas granulares. Las neuronas granulares migran desde esta capa exterior para formar una capa interna conocida como capa interna de gránulos. La capa granular externa deja de existir en el cerebelo maduro, dejando solo células granulares en la capa granular interna. La sustancia blanca cerebelosa puede ser una tercera zona germinal en el cerebelo; sin embargo, su función como zona germinal es controvertida.
Imágenes Adicionales
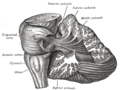
Disección que muestra las fibras de proyección del cerebelo.
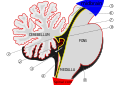
Esquema del techo del cuarto ventrículo. La flecha está en el agujero de Majendie.

Vista sagital media del cerebro humano
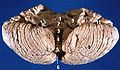
Vista anterior del cerebelo humano, con números que indican hitos destacados



